Startseite - Impressum - Datenschutzerklärung - News - Ich diene dem, der hört - https://offenunddirekt.de - Mein Gratis-Coaching für Dich - Kraft tanken - Spirituelles - Kunst - Technik - Literatur - Kultur - Politik - Architektur - Spaß - Psychologie - Spiele - Fotos + Animationen - Gästebuch - Persönliches - Blog - Die Stimme Gottes - mein Youtube Kanal - erstes Rätsel - Renaissance Forum - Chat - Quiz - Rätselverzeichnis - Wie alles begann ... - Zufallsrätsel - Zufallsspiel - letztes Rätsel
Galerie - A - B - C - D - E - F- G - H - I - J - K - L - M - N - O - P - Q - R - S - T - U - V - W - X - Y - Z |
Ernst Haeckel - Die Natürliche Schöpfungsgeschichte 1868
Sechszehnter Vortrag.
Stammbaum und Geschichte des Pflanzenreichs.
(Hirezu Taf. II.)
Meine Herren! Jeder Versuch, den wir zur Erkenntniß des Stammbaums irgend einer kleineren oder größeren Gruppe von blutsverwandten Organismen unternehmen, hat sich zunächst an das bestehende "natürliche System" dieser Gruppe anzulehnen. Denn obgleich das natürliche System der Thiere, Protisten und Pflanzen niemals endgültig festgestellt werden, vielmehr immer nur einen mehr oder weniger annähernden Grad von Erkenntniß der wahren Blutsverwandtschaft darstellen wird, so wird es nichts desto weniger jederzeit die hohe Bedeutung eines hypothetischen Stammbaums behalten. Allerdings wollen die meisten Zoologen, Protistiker und Botaniker durch ihr "natürliches System" nur im Lapidarstyl die subjectiven Anschauungen ausdrücken, die ein jeder von Ihnen von der objectiven"Formverwandtschaft" der Organismen besitzt. Allein diese Formverwandtschaft ist ja im Grunde, wie Sie gesehen haben, nur die nothwendige Folge der wahren Blutsverwandtschaft. Daher wird jeder Morphologe, welcher unsere Erkenntniß des natürlichen Systems fördert, gleichzeitig, er mag wollen oder nicht, auch unsere Erkenntniß des Stammbaums fördern. Je mehr das natürliche System seinen Namen wirklich verdient, je fester es sich auf die übereinstimmenden Resultate der vergleichenden Anatomie, Ontogenie und Paläontologie gründet, desto sicherer dürfen wir dasselbe als den annähernden Ausdruck des wahren Stammbaums betrachten.
Indem wir uns nun zu unserer heutigen Aufgabe die Genealogie des Pflanzenreichs stecken, werden wir, jenem Grundsatze gemäß, zunächst einen Blick auf das natürliche System des Pflanzenreichs zu werfen haben, wie dasselbe heutzutage von den meisten Botanikern mit mehr oder minder unbedeutenden Abänderungen angenommen wird. Danach zerfällt zunächst die ganze Masse aller Pflanzen in zwei Hauptgruppen. Diese obersten Hauptabtheilungen oder Unterreiche sind noch dieselben, welche vor mehr als einem Jahrhundert Carl Linné, der Begründer der systematischen Naturgeschichte (vergl. oben S. 32) unterschied und welche er Cryptogamen oder Geheimblühende und Phanerogamen oder Offenblühende nannte. Die letzteren theilte Linné in senem künstlichen Pflanzensystem nach der verschiedenen Zahl, Bildung und Verbindung der Staubgefäße in 23 verschiedene Klassen, und diesen fügte er dann als 24ste und letzte Klasse die Cryptogamen an.
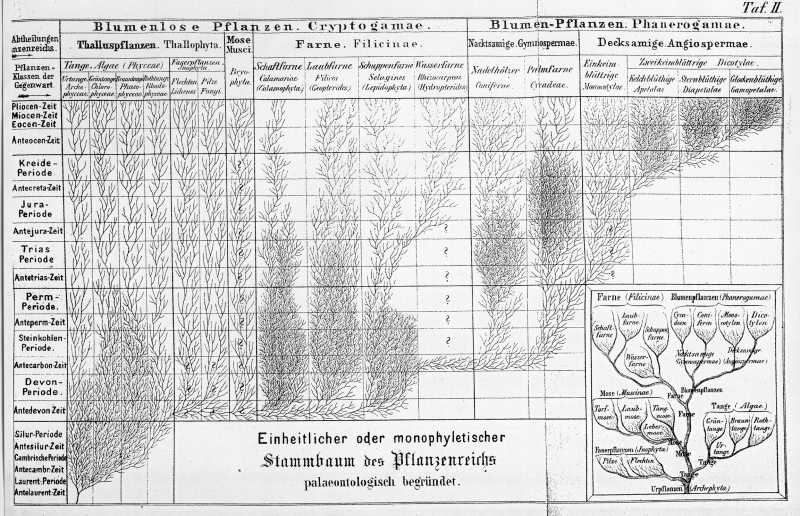
Die Cryptogamen, die geheimblühenden oder blüthenlosen Pflanzen, welche früherhin nur wenig beobachtet wurden, haben durch die eingehenden Forschungen der Neuzeit eine so große Mannichfaltigkeit der Formen, und eine so tiefe Verschiedenheit im gröberen und feineren Bau offenbart, daß wir unter denselben nicht weniger als vierzehn verschiedene Klassen unterscheiden müssen, während die Zahl der Klassen unter den Blüthenpflanzen oder Phanerogamen auf vier beschränken können. Diese achtzehn Klassen des Pflanzenreichs aber gruppieren sich naturgemäß wiederum dergestalt, daß wir im Ganzen sechs Hauptklassen (oder Kladen, d. h. Aeste) des Pflanzenreichs unterscheiden können. Zwei von diesen sechs Hauptklassen fallen auf die Blüthenpflanzen, vier dagegen auf die Blüthenlosen. Wie sich jene 18 Klassen auf diese sechs Hauptklassen, und die letztren wiederum auf die Hauptabtheilungen des Pflanzenreichs vertheilen, zeigt Ihnen übersichtlich die nachstehende Tabelle und der Stammbaum auf Taf. II.
Das Unterreich der Cryptogamen oder Blumenlosen kann man zunächst naturgemäß in zwei Hauptabtheilungen oder Stammgruppen zerlegen, welche sich in ihrem inneren Bau und in ihrer äußren Form sehr wesentlich unterscheiden, nämlich die Thalluspflanzen und die Prothalluspflanzen. Die Stammgruppe der Thalluspflanzen umfaßt die beiden großen Hauptklassen der Tange oder Algen, welche im Wasser leben, und der Faserpflanzen oder Inophyten (Flechten und Pilze), welche außerhalb des Wassers, auf der Erde, auf Steinen, Baumrinden, auf verwesenden organischen Körpern u. s. w. wachsen. Die Stammgruppe der Prothalluspflanzen dagegen enthält die beiden formenreichen Hauptklassen der Mose und Farne
Alle Thalluspflanzen oder Thallophyten sind sofort daran zu erkennen, daß man an ihrem Körper die beiden Grundorgane der übrigen Pflanzen, Stengel und Blätter, noch nicht unterscheiden kann. Vielmehr ist der ganze Leib aller Tange und aller Faserpflanzen eine aus einfachen Zellen zusammengesetzte Masse, wleche man als Laubkörper oder Thallus bezeichnet. Dieser Thallus ist noch nicht in Stengel und Blatt differenzirt. Hierdurch, sowie durch viele andere Eigenthümlichkeiten stellen sie die Thallophyten allen übrigen Pflanzen, nämlich den beiden Hauptgruppen der Prothalluspflanzen und der Blüthenpflanzen gegenüber und man hat deßhalb auch häufig die letzteren beiden als Stockpflanzen oder Cormophyten zusammengefaßt. Das Verhältniß dieser drei Stammgruppen zu einander, entsprechend jenen beiden verschiedenen Auffassungen, macht Ihnen nachstehende Uebersicht deutlich.
I. Blumenlose (Cryptogamae) A. Thalluspflanzen (Thallophyta) I. Thalluspflanzen (Thallophyta) B. Prothalluspflanzen (Prothallophyta) II. Stockpflanzen (Cormophyta)
II. Blumenpflanzen (Phanerogamae) C. Blumenpflanzen (Phanerogamae)
Die Stockpflanzen oder Cormophyten, in deren Organisation bereits der Unterschied von Stengelorganen und Blattorganen entwickelt ist, bilden gegenwärtig und schon seit sehr langer Zeit die Hauptmasse der Pflanzenwelt. Allein so war es nicht immer. Vielmehr fehlten die Stockpflanzen, und zwar nicht allein die Blumenpflanzen, sondern auch die Prothalluspflanzen, noch gänzlich während jenes unermeßlich langen Zeitraums, welcher als das archolithische oder primordiale Zeitalter den Beginn und den ersten Hauptabschnitt der organischen Erdgeschichte bildet. Sie erinnern sich, daß während dieses Zeitraums sich die laurentischen, cambrischen und silurischen Schichtensysteme ablagerten, deren Dicke zusammengenommen ungefähr 70,000 Fuß beträgt. Da nun die Dicke aller darüber liegenden jüngeren Schichten, von den devonischen bis zu den Ablagerungen der Gegenwart zusammen nur ungefähr 60,000 Fuß erreicht, so konnten wir hieraus allein den auch aus anderen Gründen wahrscheinlichen Schluß ziehen, daß jenes archolitische oder primordiale Zeitalter eine längere Dauer besaß, als die ganze darauf folgende Zeit bis zur Gegnwart. Während dieses ganzen unermeßlichen Zeitraums, der vielleicht viele Millionen von Jahrhunderten umschloß, scheint das Pflanzenleben auf unserer Erde ausschließlich durch die Stammgruppe der Thalluspflanzen, und zwar nur durch die Hauptklasse der wasserbewohnenden Thalluspflanzen, durch die Tange und Algen, vertreten gewesen zu sein. Wenigstens gehören alle versteinerten Pflanzenreste, welche wir mit Sicherheit aus der Primordialzeit kennen, ausschließlich dieser Hauptklasse an.

{ Abbildung von Seite 352}
Da auch alle Thierreste dieses ungeheuren Zeitraums nur wasserbewohnenden Thieren angehören, so schließen wir daraus, daß landbewohnende Organismen damals noch gar nicht existirten.
Schon aus diesen Gründen muß die erste und unvollkommenste Hauptklasse des Pflanzenreichs, die Abtheilung der Tange oder Algen für uns von ganz besonderer Bedeutung sei. Dazu kommt noch das hohe Interesse, welches uns diese Hauptklasse, auch an sich betrachtet, gewährt. Trotz ihrer höchst einfachen Zusammensetzung aus gleichartigen oder nur wenig differenzirten Zellen zeigen die Tange dennoch eine außerordentliche Mannichfaltigkeit verschiedener Formen.
Einerseits gehören dazu die einfachsten und unvollkommensten aller Gewächse, andrerseits sehr entwickelte und eigenthümliche Gestalten. Ebenso wie in der Vollkommenehit und Mannichfaltigkeit ihrer äußeren Formbildung unterscheiden sich die verschiedenen Algengruppen auch in der Körpergröße. Auf der tiefsten Stufe finden wir die winzig kleinen Protococcus-Arten, von denen mehrere Hunderttausend auf den Raum einen Stecknadelkopfes gehen. Auf der höchsten Stufe bewundern wir in den riesenmäßigen Makrocysten, welche eine Länge von 300-400 Fuß erreichen, die längsten von allen Gestalten des Pflanzenreichs. Und wenn nicht aus diesen Gründen, so müßten die Algen schon deßhalb unsere besondere Aufmerksamheit erregen, weil sie die Anfänge des Pflanzenlebens bilden und die Stammformen aller übrigen Pflanzengruppen enthalten, vorausgesetzt daß unsere Hypothese von einem gemeinsamen Ursprung aller Pflanzengruppen richtig ist (Taf. II).
Leider werden die Meisten von Ihnen sich nur eine sehr unvollkommene Vorstellung von dieser höchst interessanten Hauptklasse des Pflanzenreichs machen können, weil Sie davon nur die verhältnißmäßig kleinen und einfachen Vertreter kennen werden, welche das süße Wasser bewohnen. Die schleimigen grünen Wasserfäden und Wasserflocken in unseren Teichen und Brunnentrogen, die hellgrünen Schleimüberzüge auf allerlei Holzwerk, welches längere Zeit mit Wasser in Berührung war, die gelbgrünen schaumigen Schleimdecken auf den Tümpeln unserer Dörfer, die grünen Haarbüscheln gleichenden Fadenmassen, welche überall im stehenden und fließenden Süßwasser vorkommen, sind größtentheils aus verschiedenen Tangarten zusammengesetzt. Aber nur diejenigen von Ihnen, welche die Meeresküste besucht haben, welche an den Küsten von Helgoland und von Schleswig-Holstein die ungeheuren Massen des ausgeworfenen Seetangs bewundert, ober an den Felsenufern des Mittelmeeres die zierlich gestaltete und lebhaft gefärbte Tangvegetation auf dem Meeresboden selbst durch die klare blaue Fluth hindurch erblickt haben, wissen die Bedeutung der Tangklasse annähernd zu würdigen. Und dennoch geben selbst diese formenreichen untermeerischen Algenwälder der europäischen Küsten nur eine schwache vorstellung von den colossalen Sargassowäldern des atlantischen Oceans, jenen ungeheuren Tangbänken, welche einen Flächenraum von ungefähr 4000 Quadratmeilen bedecken, und welche dem Columbus auf seiner Entdeckungsreise die Nähe des Festlandes vorspiegelten.
Aehnliche, aber weit ausgedehntere Tangwälder wuchsen in dem primoridalen Urmeere wahrscheinlich in dichten Massen, und wie zahllose Generationen dieser archolithischen Tange über einander hinstarben, bezeugen unter Anderen die mächtigen silurischen Alaunschiefer Schwedens, deren eigenthümliche Zusammensetzung wesentlich von jenen untermeerischen Algenmassen herrührt.
Wir unterscheiden in der Hauptklasse der Tange oder Algen vier verschiedene Klassen, deren jede wiederum in mehrere Ordnungen und Familien zerfällt. Diese ihrerseits enthalten wieder eine große Menge verschiedener Gattungen und Arten. Wir bezeichnen diese vier Klassen als Urtange oder Archephyceen, Grüntange oder Chlorophyceen, Brauntange oder Phaeophyceen, und Rothtange oder Rhodophyceen.
Die erste Klasse der Tange, die Urtange (Archephyceae) könnten auch Urpflanzen (Archephyta) genannt werden, weil dieselben die einfachsten und unvollkommensten von allen Pflanzen enthalten, und insbesondere jene ältesten aller pflanzlichen Organismen, welche allen übrigen Pflanzen den Ursprung gegeben haben. Es gehören hierher also zunächst jene allerältesten vegetabilischen Moneren, welche im Beginne der antelaurentischen Periode durch Urzeugung entstanden sind. Ferner müssen wir dahin alle jene Pflanzenformen einfachster Organisation rechnen, welche aus jenen sich zunächst in antelaurentischer Zeit entwickelt haben, und welche den Formwerth einer einzigen Plastide besaßen. Zunächst waren dies solche Urpflänzchen, deren ganzer Körper eine einfache Cytode (eine kernlose Plastide) bildete, und weiterhin solche, die bereits durch Sonderung eines Kernes um Plasma den höheren Formwerth einer einfachen Zelle erreicht hatten (Vergl. oben S. 285).
Noch in der Gegenwarth leben verschiedene einfachste Tangarten, welche von diesen ursprünglichen Urpflanzenformen sich nur wenig entfernt haben. Dahin gehört eine große Anzahl von höchst einfachen, meist mikroskopisch kleinen Pflänzchen, deren ganzer Körper noch heutzutage in vollkommen ausgebildeten Zustande nur den Formwerth einer einfachen Plastide, einweder einer Cytode oder einer Zelle besitzt; oder bei denen nur eine geringe Anzahl von einfachen und gleichartigen Zellen zur Bildung des Thalluskörpers zusammentritt. Die Tangfamilien der Codiolaceen, Protococcaceen, Desmidiaceen, Palmellaceen und einige andere würden hierher zu rechnen sein. Auch die merkwürdige Gruppe der Phycochromaceen (Chroococcaceen und Oscillarineen) würde man hierher ziehen können, falls man diese nicht lieber als einen selbstständigen Stamm des Protistenreichs ansehen will (Vergl. oben S. 328). Endlich würde man zu den Urtangen auch jene außerordentlich merkwürdigen Schlauchalgen oder Siphoneen rechnen können, deren Körper bei ansehnlicher Größe und sehr entwickelter äußerer Form dennoch aus einer einzigen einfachen Plastide besteht. Manche von diesen Siphoneen erreichen eine Größe von mehreren Füßen und gleichen einem zierlichen Mose (Bryopsis) oder einem Bärlappe oder gar einer vollkommenen Blüthenpflanze mit Stengen, Wurzeln und Blättern (Caulerpa). Und dennoch besteht dieser ganze große und vielfach äußerlich differenzirte Körper innerlich aus einem ganz einfachen Schlauche, der nur den Formwerth einer einzigen Cytode besitzt. Diese wunderbaren Siphoneen, Bryopsen und Caulerpen, zeigen uns, wie weit es die einzelne Plastide als ein einfachstes Individuum erster Ornung durch fortgesetzte Anpassung an die Verhältnisse der Außenwelt bringen kann. Es ist sehr wahrscheinlich, daß ähnliche Urpflanzen, deren weicher Körper aber nicht der fossilen Erhaltung fähig war, in großer Masse und Mannichfaltigkeit das antelaurentische Urmeer bevölkerten und einen großen Formenreichthum entfalteten, ohne doch die Individualitätsstufe einer einfachen Plastide zu überschreiten.
An die Urpflanzen oder Urtange schließt sich als zweite Klasse der Algen zunächst die Gruppe der Grüntange (Chlorophyceae) oder Grünalgen (Chloralgae) an. Gleich der Mehrzahl der ersteren sind auch sämmliche Grüntange grün gefäbt, und zwar durch denselben Farbstoff, das Blattgrün oder Chlorophyll, welches auch die Blätter aller höheren Gewächse grün färbt. Zu dieser Klasse gehören außer einer großen Anzahl von niederen Seetangen die allermeisten Tange des süßen Wassers, die gemeinen Wasserfäden oder Conferven, die grünen Schleimkugeln oder Glöosphären, der hellgrüne Wassersalat oder die Ulven, welche einem sehr dünnen und langen Salatblatte gleichen, ferner zahlreiche mikroskopisch kleine Tange, welche in dichter Masse zusammengehäuft einen hellgrünen schleimigen Ueberzug über allerlei im Wasser liegende Gegenstände, Holz, Steine u. s. w. bilden, sich aber durch die zellige Zusammensetzung und Sonderung ihres Körpers bereits weit über die Urtange erheben. Da die Grüntange, gleich den Urtangen, meistens einen sehr weichen Körper besitzen, waren sie nur sehr selten der Versteinerung fähig. Es kann aber wohl nicht bezweifelt werden, daß auch diese Algenklasse, welche sich zunächst aus der vorhergehenden entwickelt hat, gleich jener in früherer Zeit die süßen und salzigen Gewässer der Erde in sehr viel größerer Ausdehnung und Mannichfaltigkeit bevölkerte.
In der dritten Klasse, derjenigen der Brauntange (Phaeophyceae) oder Schwarztange (Fucoideae) erreicht die Hauptklasse der Algen ihren höchsten Entwickelungsgrad, wenigstens in Bezug auf die körperliche Größe. Die charakteristische Farbe der Fucoideen ist meist ein mehr oder minder dunkles Braun, bald mehr in Olivengrün und Gelbgrün, bald mehr in Braunroth und Schwarz übergehend. Hierher gehören die größten aller Tange, welche zugleich die längsten von allen Pflanzen sind, die colossalen Riesentange, unter denen Macrocystis pyrifera) an der californischen Küste eine Länge von 400 Fuß erreicht. Aber auch unter unseren einheimischen Tangen gehören die ansehnlichsten Formen zu dieser Gruppe, so namentlich der stattliche Riementang (Laminaria), dessen schleimige olivengrüne Thalluskörper, riesigen Blättern von 10-15 Fuß Länge, 1/2 - 1 Fuß Breite gleichend, in großen Massen an der Küste der Nord- und Ostseen ausgeworfen werden. Auch der in unseren Meeren gemeine Blasentang (Fucus vesiculosus), dessen mehrfach gabelförmig gespaltenes Laub durch viele eingeschlossenen Luftblasen, (wie bei vielen anderen Brauntangen) auf dem Wasser schwimmend erhalten wird, gehört zu dieser Klasse; ebenso der freischwimmende Sargassotang (Sargassum bacciferum), welcher die schwimmenden Wiesen oder Bänke des Sargassomeeres bildet. Aehnliche Brauntange sind es wahrscheinlich auch zum größten Theile gewesen, welche während der Primordialzeit die charakteristischen Tangwälder dieses endlosen Zeitraums zusammengesetzt haben. Die versteinerten Reste, welche uns von denselben (vorzüglich aus der silurischen Zeit) erhalten sind, können uns allerdings nur eine schwache Vorstellung davon geben, weil auch diese Tange, gleich den meisten anderen, sich nur schlecht zur Erhaltung im fossilen Zustande eignen.
Weniger bedeutend war damals vielleicht die vierte und letzte Klasse der Tange, diejenige der Rosentange (Rhodophyceae) oder Rothtange (Florideae). Zwar entfaltet auch diese Klasse einen großen Reichthum verschiedener Formen. Allein die meisten derselben sind von viel geringerer Größe als die Brauntange. Uebrigens stehen sie den letzteren an Vollkommenheit und Differenzirung der äußeren Form keineswegs nach, übertreffen dieselben vielmehr in mancher Beziehung. Hierher gehören die schönsten und zierlichsten aller Tange, welche sowohl durch die feine Fiederung und Zertheilung ihres Laubkörpers, wie durch reine und zarte rothe Färbung zu den reizendsten Pflanzen gehören. Die charakteristische rothe Farbe ist bald ein tiefes Purpur-, bald ein brennendes Scharlach-, bald ein zartes Rosenroth, und geht einerseits in violette und purpurblaue, andrerseits in braune und grüne Tinten in bewunderungswürdiger Pracht über. Wer von Ihnen eines unserer nordischen Seebäder besucht hat, wird gewiß schon mit Staunen die reizendsten Formen dieser Florideen betrachtet haben, welche auf weißem Papier, zierlich angetrocknet, vielfach zum Verkaufe angeboten werden. Die meisten Rothtange sind leider so zart, daß sie gar nicht der Versteinerung fähig sind, so die prachtvollen Ptiloten, Plokamien, Delesserien u. s. w. Doch giebt es einzelne Formen, die Chondrien und Sphärokokken, welche einen härteren, oft fast knorpelhaften Thallus besitzen, und von diesen sind uns auch manche versteinerte Reste, namentlich aus silurischen, devonischen und Kohleschichten, sspäter besonders aus dem Jura erhalten worden. Wahrscheinlich nahm auch diese Klasse an der Zusammensetzung der archolithischen Tangflora wesentlichen Antheil.
Wenn Sie nun nochmals einen Rückblick auf die Flora der Primordialzeit werfen, welche ausschließlich von der Hauptklasse der Tange gebildet wurde, so finden Sie, daß die vier untergeordneten Klassen derselben wahrscheinlich in ähnlicher Weise an der Zusammensetzung jener submarinen Wälder des Urmeeres sich betheiligt haben, wie in der Gegenwart die vier physiognomischen Vegetationstypen der stämmigen Bäume, der blumigen Kräuter, des buschigen Grases und der zartlaubigen Farne und Mose an der Zusammensetzung unserer Landwälder Theil nehmen. Man könnte in dieser Beziehung sagen, daß die unterseeischen Waldbäume der Primordialzeit durch die mächtigen Brauntange oder Fucoideen gebildet wurden. Die farbigen Blumen zu den Füßen dieser Baumriesen wurden durch die bunten Rothtange oder Florideen vertreten. Das grüne Gras dazwischen bildeten die haarbüscheligen Grüntange oder Chloralgen. Das zarte Laub der Farne und Mose endlich, welches den Boden unserer Wälder bedeckt, die Lücken ausfüllt, welche die anderen Pflanzen übrig lassen, und selbst auf den Stämmen der Bäume sich ansiedelt, wird damals ähnliche Vertreter in den mosähnlichen und farnähnlichen Siphoneen, in den Caulerpen und Bryopsen aus der Klasse der Urtange oder Archephyten gehabt haben.
Was die Verwandtschaftsverhältnisse der verschiedenen Tangklassen zu einander und zu den übrigen Pflanzen betrifft, so bilden höchst wahrscheinlich, wie schon bemerkt, die Urtange oder Archephyten die gemeinsame Wurzel des Stammbaums, nicht allein für die verschiedenen Tangklassen, sondern für das ganze Pflanzenreich. Aus den nackten vegetabilischen Moneren, welche sich im Beginn der antelaurentischen Periode entwickelten, werden zunächst Hüllcytoden entstanden sein (S. 286), indem der nackte, strukturlose Eiweißleib der Moneren sich an der Oberfläche krustenartig verdichtete oder eine Hülle ausschwitzte.
Späterhin werden dann aus diesen Hüllcytoden echte Pflanzenzellen geworden sein, indem im Innerern sich ein Kern oder Nucleus von dem umgebenden Zellstoff oder Plasma sonderte. Die drei Klassen der Grüntange, Brauntange und Rothtange sind wahrscheinlich drei gesonderte Stämme, welche unabhängig von einander aus der gemeinsamen Wurzelgruppe der Urtange entstanden sind und sich dann (ein jeder in seiner Art) weiter entwickelt und vielfach in Ordnungen und Familien verzweigt haben. Die Brauntange und Rothtange haben keine weitere Blutsverwandtschaft zu den übrigen Klassen des Pflanzenreichs. Diese letzteren sind vielmehr aus den Urtangen entstanden, und zwar direkt oder durch Vermittlung der Grüntange. Wahrscheinlich sind einerseits die Mose und Farne, andrerseits die Flechten und Pilze unabhängig von einander aus den Urtangen entstanden, die ersteren vielleicht durch Vermittlung der Grüntange. Die Blumenpflanzen oder Phanerogamen haben sich jedenfalls erst später aus den Farnen entwickelt.
Als zweite Hauptklasse des Pflanzenreichs haben wir oben die Faserpflanzen (Inophyta) angeführt. Wir verstanden darunter die beiden naheverwandten Klassen der Flechten und Pilze. Es ist möglich, daß diese Thalluspflanzen nicht aus den Urtangen entstanden sind, sondern aus einer oder mehreren Moneren, die unabhängig von letzteren durch Urzeugung entstanden. Auch ist noch der andere Fall denkbar, daß die verschiedenen Ordnungen sowohl der Flechtenklasse als der Pilzklasse, und namentlich die niedersten Formen beider Klassen, einer größeren Anzahl von verschiedenen archigonen (d. h. durch Urzeugung entstandenen) Moneren ihren Ursprung verdanken. Jedenfalls sind beide Klassen nicht als Stammeltern der höheren Pflanzenklassen zu betrachten. Sowohl die Flechten als die Pilze unterscheiden sich von diesen durch die Zusammensetzung ihres weichen Körpers aus einem dichten Geflecht von sehr langen, vielfach verschlungenen eigenthümlichen Fadenzellen oder Fasern, weshalb wir sie eben in der Hauptklasse der Faserpflanzen zusammenfassen. Irgend bedeutende fossilen Reste konnten dieselben wegen ihrer eigenthümlichen Beschaffenheit nicht hinterlassen, und so müssen wir denn die paläontologische Bedeutung und Entwickelung derselben mehr erraten, als daß wir sie mit Sicherheit aus Petrefacten erkennen könnten.
Die Klasse der Flechten (Lichenes) hat wahrscheinlich zu allen Zeiten dieselbe äußerlich untergeordnete Rolle gespielt, wie in der Gegenwart. Die meisten Flechten bilden mehr oder weniger unansehnliche, formlose oder unregelmäßig zerrissene, krustenartige Ueberzüge auf Steinen, Baumrinden u. s. w. Die Farbe derselben wechselt in allen möglichen Abstufungen vom reinsten Weiß, durch Gelb, Roth, Grün, Braun, bis zum dunkelsten Schwarz. Wichtig sind indessen viele Flechten in der Oekonomie der Natur dadurch, daß sie sich auf den trockensten und unfruchtbarsten Orten, insbesondere auf dem nackten Gestein ansiedeln können, auf welchem keine andere Pflanze leben kann. Die harte schwarze Lava, welche in vulkanischen Gegenden viele Quadratmeilen Bodens bedeckt, und welche oft Jahrhunderte lang jeder Pflanzenansiedelung den hartnäckigsten Widerstand leistet, wird zuerst immer von Flechten bewältigt. Weiße oder graue Steinflechten (Stereocaulon) sind es, welche auf den ödesten und todtesten Lavafeldern immer mit der Urbarmachung des nackten Felsenbodens beginnen und denselben für die nachfolgende höhere Vegetation erobern. Ihre absterbenden Leiber bilden die erste Dammerde, in welcher nachher Mose, Farne und Blüthenpflanzen festen Fuß fassen können. Auch gegen klimatische Unbilden sind die zähen Flechten unempfindlicher als alle anderen Pflanzen. Daher überziehen ihre trockenen Krusten die nackten Felsen noch in den höchsten, größtentheils mit ewigem Schnee bedeckten Gebirgshöhen, in denen keine anderen Pflanze mehr ausdauern kann. Dürfen wir aus diesen Lebenseigenthümlichkeiten der Flechten auf ihre geschichtliche Entwickelung und Bedeutung schließen, so ist es sehr wahrscheinlich, daß Flechten die ersten landbewohnenden Pflanzen waren. Aller Wahrscheinlichkeit nach entstanden die ersten Flechten im Beginn des primären Zeitalters, im Anfang der antedevonischen Zeit, dadurch, daß einzelne Urtange oder Archphyten von ihrer ursprünglichen Geburtsstätte, dem primordialen Urmeere, auf das eben geborene antedevonische Festland, auf die ersten Erhebungen der festen Erdrinde über den Spiegel des silurischen Meeres übersiedelten. Indem so die Flechten die nackte Oberfläche der ersten Festlandsfelsen für die nachfolgenden Mose und Farne eroberten, gewannen sie eine paläontologische Bedeutung, auf welche wir aus den dürftigen versteinerten Bruchstücken derselben, und aus ihrem unansehnlichen Aeußeren keineswegs schließen könnten.
Die zweite Klasse der Faserpflanzen, die Pilze (Fungi) werden irrthümlich oft Schwämme genannt und daher mit den echten thierischen Schwämmen oder Spongien verwechselt. Sie zeigen einerseits so viele Verwandtschaftsbeziehungen zu den Flechten und sind durch so viele Uebergangsformen (namentlich die Kernschwämme oder Pyrenomyceten) mit denselben verbunden, daß man beide Klassen kaum trennen kann, und es für das Natürlichste halten dürfte, eine Abstammung der Pilze von den Flechten anzunehmen. Andrerseits aber haben die meisten Pilze so viel Eigenthümliches und weichen namentlich durch ihre eigenthümliche Ernährungsweise so sehr von allen übrigen Pflanzen ab, daß man sie als eine ganz besondere Hauptgruppe des Pflanzenreichs betrachten könnte. Die übrigen Pflanzen leben größtentheils von anorganischer Nahrung, d. h. von einfachen und festen Kohlenstoffverbindungen, welche sie zu verwickelteren zusammensetzen. Sie athmen Kohlensäure ein und Sauerstoff aus. Die Pilze dagegen leben größtentheils, gleich den Thieren, von organischer Nahrung, d. h. von verwickelten und lockeren Kohlenstoffverbindungen, welche sie zersetzen. Sie athmen Sauerstoff ein und Kohlensäure aus, wie die Thiere. Auch bilden sie niemals das Blattgrün oder Chlorophyll, welches für die meisten übrigen Pflanzen zu charakteristisch ist.
Daher haben schon wiederholt hervorragende Botaniker den Vorschlag gemacht, die Pilze ganz aus dem Pflanzenreiche zu entfernen und als ein besonderes drittes Reich zwischen Thier- und Pflanzenreich zu setzen. Dadurch würde unser Protistenreich einen sehr bedeutenden Zuwachs erhalten, und ich habe kürzlich in einer neuen Begrenzung des Protistenreichs die Pilze in der That als eine besondere Protistenklasse neben die Phycochromaceen und die Schleimpilze (Myxomyceten) gestellt15). Da jedoch die meisten von Ihnen wohl mehr geneigt sein werden, der herkömmlichen Anschauung gemäß die Pilze als echte Pflanzen zu betrachten, lasse ich sie hier im Pflanzenreiche stehen, und verbinde sie mit den Flechten, denen sie im anatomischen inneren Bau am nächsten verwandt sind. Ob dieselben aber aus den Flechten oder aus den Urtangen entstanden sind, oder ob sie, was mir das Wahrscheinlichste ist, mehreren selbstständigen archigonen Moneren ihren Ursprung verdanken, das will ich hier ganz dahingestellt sein lassen.
Indem wir nun die Pilze, Flechten und Tange, welche gewöhnlich als Thalluspflanzen zusammengefaßt werden, verlassen, betreten wir das Gebiet der zweiten großen Hauptabtheilung des Pflanzenreichs, der Prothalluspflanzen (Prothallophyta), welche von anderen als phyllogonische Kryptogamen bezeichnet werden (im Gegensatz zu den Thalluspflanzen oder thallogonischen Kryptogamen). Dieses Gebiet umfaßt die beiden Hauptklassen der Mose und Farne. Hier begegnen wir bereits allgemein (wenige der untersten Stufen ausgenommen) der Sonderung des Pflanzenkörpers in zwei verschiedene Grundorgane: Stengel oder Axenorgane, und Blätter oder Seitenorgane. Hierin gleichen die Prothalluspflanzen bereits den Blumenpflanzen und daher faßt man sie neuerdings auch häufig mit diesen als Stockpflanzen oder Cormophyten zusammen. Andrerseits gleichen die Mose und Farne den Thalluspflanzen durch den Mangel einer echten Blüthe oder Blume, und daher stellte sie schon Linné mit diesen als Kryptogamen zusammen, im Gegensatz zu den Blumenpflanzen oder Phanerogamen.
Unter dem Namen "Prothalluspflanzen" vereinigen wir die nächstverwandten Mose und Farne deshalb, weil bei Beiden sich ein sehr eigenthümlicher und charakteristischen Generationswechsel in der individuellen Entwickelung findet. Jede Art nämlich tritt in zwei verschiedenen Generationen auf, von denen man die eine gewöhnlich als Vorkeim oder Prothallium bezeichnet, die andere dagegen als den eigentlichen Stock oder Cormus des Moses oder des Farns betrachtete. Die erste und ursprüngliche Generation, der Vorkeim oder Prothallus, auch Prothallium genannt, steht noch auf jener niederen Stufe der Formbildung, welche alle Thalluspflanzen zeitlebens zeigen, d. h. es sind Stengel und Blattorgane noch nicht gesondert, und der ganze zellige Körper des Vorkeims stellt einen einfachen Thallus dar. Die zweite und vollkommenere Generation der Mose und Farne dagegen, der Stock oder Cormus, bildet einen viel höher organisirten Körper, welcher wie bei den Blumenpflanzen in Stengel und Blatt gesondert ist, ausgenommen bei den niedersten Mosen, bei welchen auch diese Generation noch auf der niederen Stufe der ursprünglichen Thallusbildung stehen bleibt. Mit Ausnahme dieser letzteren erzeugt allgemein bei den Mosen und Farnen die erste Generation, der thallusförmige Vorkeim, eine stockförmige zweite Generation mit Stengel und Blättern; diese erzeugt wiederum den Thallus der ersten Generation u. s. w. Es ist also, wie bei dem gewöhnlichen einfachen Generationswechsel der Thiere, die erste Generation der dritten, fünften u. s. w., die zweite dagegen der vierten, sechsten u. s. w. gleich. (Vergl. oben S. 161).
Von den beiden Hauptklassen der Prothalluspflanzen stehen die Mose im Allgemeinen auf einer viel tieferen Stufe der Ausbildung, als die Farne und vermitteln namentlich in anatomischer Beziehung den Uebergang von den Thalluspflanzen und speicell von den Tangen zu den Farnen. Ob jedoch dadurch ein genealogischer Zusammenhang der Mose und Farne angedeutet wird, ist noch zweifelhaft. Jedenfalls sind die Mose direkt aus Thalluspflanzen und zwar wahrscheinlich entweder aus Grüntangen oder aus Urtangen entstanden. Die Farne stammen entweder in gleicher Weise, als ein von den Mosen unabhängiger Stamm, von den Thalluspflanzen ab, oder sie haben sich aus unbekannten ausgestorbenen Mosformen entwickelt. Für die Schöpfungsgeschichte sind die Farne von weit höherer Bedeutung als die Mose.
Die Hauptklasse der Mose (Muscinae, auch Musci oder Bryophyta genannt) enthält die niederen und unvollkommneren Pflanzen der Prothallophytengruppe, welche sich zunächst an die Thalluspflanzen anschließen. Meistens ist ihr Körper so zart und vergänglich, daß er sich nur schlecht zur kenntlichen Erhaltung in versteinertem Zustande eignet. Daher sind die fossilen Reste von allen Mosklassen selten und unbedeutend. Die meisten deutlich erhaltenen stammen aus den tertiären Gesteinen. Jedoch haben zweifelsohne die Mose schon in viel früherer Zeit sich aus den Thalluspflanzen, vermuthlich aus den Urtangen oder Grüntangen entwickelt. Wasserbewohnende Uebergangsformen von letzteren zu den Mosen gab es wahrscheinlich schon in der Primordialzeit und landbewohnende in der Primärzeit. Die Mose der Gegenwart, aus deren stufenweis verschiedener Ausbildung die vergleichende Anatomie Einiges auf ihre Genealogie schließen kann, zerfallen in vier verschiedene Klassen, nämlich 1. die Tangmose; 2. die Lebermose; 3. die Laubmose und 4. die Torfmose.
Auf der tiefsten Stufe der mosartigen Pflanzen steht die erste Klasse, die Tangmose (Characeae oder Charobrya). Hierher gehören die tangartigen Armleuchterpflanzen (Chara) und Glanzmose (Nitella), welche mit ihren grünen fadenförmigen, quirlartigen von gabelspaltigen Aesten umstellten Stengeln in unseren Teichen und Tümpeln oft dichte Bänke bilden. Einerseits nähern sich die Characeen im anatomischen Bau, bsonders der Fortpflanzungsorgane, den Mosen und werden diesen neuerdings unmittelbar angereiht. Andrerseits stehen sie durch viele Eigenschaften tief unter den übrigen Mosen und schließen sich vielmehr den Grüntangen oder Chlorophyceen an. Man könnte sie daher wohl als übrig gebliebene und eigenthümlich ausgebildete Abkömmlinge von jenen Grüntangen betrachten, aus denen sich die übrigen Mose entwickelt haben. Durch manche Eigenthümlichkeiten sind übrigens die Tangmose so sehr von allen übrigen Pflanzen verschieden, daß viele Botaniker sie als eine besondere Hauptabtheilung des Pflanzenreichs betrachten. Man könnte sogar daran denken, daß sie einen ganz besonderen Stamm bilden, welcher sich selbstständig aus einer eigenen archigonen Monerenform entwickelt hat. Die Versteinerungskunde kann uns darüber nicht belehren. Die zweite Klasse der Mose bilden die Lebermose (Hepaticae oder Thallobrya). Die hierher gehörigen Mose sind meistens wenig bekannte, kleine und unansehnliche Formen. Die niedersten Formen derselben besitzen noch in beiden Generationen einen einfachen Thallus, wie die Thalluspflanzen, so z. B. die Riccien und Marchantien. Die höheren Lebermose dagegen, so z. B. die Jungermannien und Verwandte, beginnen allmählich Stengel und Blatt zu sondern, und die höchsten schließen sich unmittelbar an die Laubmose an. Die Lebermose zeigen durch diese Uebergangsbildung ihre direkte Abstammung von den Thallophyten, und zwar wahrscheinlich von den Grüntangen.
Diejenigen Mose, welche der Laie gewöhnlich allein kennt, und welche auch in der That den hauptsächlichsten Bestandtheil der ganzen Hauptklasse bilden, gehören zu der dritten Klasse, den Laubmosen (Musci frondosi, Musci im engeren Sinne oder Phyllobrya genannt). Hierher gehören die meisten jener zierlichen Pflänzchen, die zu dichten Gruppen vereinigt, den seidenglänzenden Mosteppich unserer Wälder bilden, oder auch in Gemeinschaft mit Lebermosen und Flechten die Rinde der Bäume überziehen. Als die Wasserbehälter, welche die Feuchtigkeit sorgfältig aufbewahren, sind sie für die Oekonomie der Natur von der größten Wichtigkeit.
Wo der Mensch schonungslos die Wälder abgeholzt und ausgerodet hat, da verschwinden mit den Bäumen auch die Laubmose, welche ihre Rinde bedeckten oder im Schutze ihres Schattens den Boden bekleideten und die Lücken zwischen den größeren Gewächsen ausfüllten. Mit den Laubmosen verschwinden aber auch die nützlichen Wasserbehälter, welche Regen und Thau sammelten und für die Zeiten der Trockniß aufgewahrten. Es entsteht dadurch eine trostlose Dürre des Bodens, welche das Aufkommen jeder ergiebigen Vegetation vereitelt. In dem größten Theile Südeuropas, in Griechenland, Italien, Sicilien, Spanien sind durch die rücksichtslose Ausrodung der Wälder die Mose vernichtet und dadurch der Boden seiner nützlichsten Feuchtigskeitsvorräthe beraubt worden; die vormals blühendsten und üppigsten Landstriche sind in dürre, öde Wüsten verwandelt. Leider nimmt auch in Deutschland neuerdings diese rohe Barbarei immer mehr überhand. Wahrscheinlich haben die kleinen Laubmose jene außerordentlich wichtige Rolle schon seit sehr langer Zeit, vielleicht seit Beginn der Primärzeit gespielt. Da aber ihre zarten Leiber ebenso wenig wie die der übrigen Mose für die deutliche Erhaltung im fossilen Zustande geeignet sind, so kann uns auch hierüber die Paläontologie keine Auskunft geben.
Als einen besonderen Zweig der Laubmosklasse haben wir endlich die vierte und letzte Mosklasse zu betrachten, die Torfmose (Sphagnaceae oder Sphagnobrya). Wahrscheinlich haben sich dieselben aus einer Abtheilung der Laubmose, vielleicht aber auch direkt aus den Lebermosen entwickelt. Auch von dieser Klasse verräth uns die Versteinerungskunde nicht den Zeitpunkt ihrer Entstehung. Auch diese Mose sind trotz ihres unscheinbaren Aeußeren doch durch ihr massenhaftes Wachsthum für den Naturhaushalt von größter Wichtigkeit. Indem ihre abgestorbenen Leiber auf dem Sumpf- und Moorboden, in dem sie wachsen, sich in vielen Generationen über einander häufen, bilden sie den Torf, der für die Bodenbildung vieler Gegenden von höchster Bedeutung ist.
Weit mehr als von den Mosen wissen wir durch die Versteinerungskunde von der außerordentlichen Bedeutung, welche die zweite Hauptklasse der Prothalluspflanzen, die der Farne, für die Geschichte der Pflanzenwelt gehabt hat. Die Farne, oder genauer ausgedrückt, die "farnartigen Pflanzen" (Filicinae oder Pteridoidae, auch Pteridophyta genannt) bildeten während eines außerordentlich langen Zeitraums, nämlich während des ganzen primären oder paläolithischen Zeitalters, die Hauptmasse der Pflanzenwelt, so daß wir dasselbe gradezu als das Zeitalter der Farnwälder bezeichnen konnten. Von Anbeginn der antedevonischen Zeit, in welcher zum ersten Male die landbewohnenden Organismen auftraten, während der Ablagerung der devonischen, carbonischen und permischen Schichten, sowie während der langen Zwischenräume zwischen den Bildungszeiten dieser Schichtensysteme, überwogen die farnartigen Pflanzen so sehr alle übrigen, daß jene Benennung dieses Zeitalters in der That gerechtfertigt ist. In den devonischen, carbonischen und permischen Schichtensystemen, vor allen aber in den ungeheuer mächtigen Steinkohlenflötzen der carbonischen oder Steinkohlenzeit, finden wir so zahlreiche und zum Theil wohl erhaltene Reste von Farnen, daß wir uns daraus ein ziemlich lebendiges Bild von der ganz eigenthümlichen Landflora des paläolithischen Zeitalters machen können. Im Jahre 1855 betrug die Gesammtzahl der damals beaknnten paläolithischen Pflanzenarten ungefähr Eintausend, und unter diesen befanden sich nicht weniger als 872 farnartige Pflanzen. Unter den übrigen 128 Arten befanden sich 77 Gymnospermen (Nadelhölzer und Palmfarne), 40 Thalluspflanzen (gröptentheils Tange) und gegen 20 nicht sicher bestimmbare Cormophyten.
Wie schon vorher bemerkt, haben sich die Farne entweder aus niederen unbekannten Mosen oder unabhängig von diesen direkt aus Thalluspflanzen, und zwar aus Grüntangen entwickelt. Wahrscheinlich fällt dieser Entwickelungsprozeß, wie der der Mose, in den Beginn der Primärzeit, in die antedevonische Zeit. In ihrer Organisation erheben sich die Farne bereits bedeutend über die Mose und schließen sich in ihren höheren Formen schon an die Blumenpflanzen an. Während bei den Mosen noch ebenso wie bei den Thalluspflanzen der ganze Körper aus ziemlich gleichartigen, wenig oder nicht differenzirten Zellen zusammengesetzt ist, entwickeln sich im Gewebe der Farne bereits jene eigenthümlich differenzirten Zellenstränge, welche man als Pflanzengefäße und Gefäßbündel bezeichnet, und welche auch bei den Blumenpflanzen vorkommen. Daher vereinigt man wohl auch die Farne als "Gefäßkryptogamen" mit den Phanerogamen, und stellt diese "Gefäßpflanzen" den "Zellenpflanzen" gegenüber, d. h. den "Zellenkryptogamen" (Mosen und Thalluspflanzen). Dieser hochwichtige Fortschritt in der Pflanzenorganisation, die Bildung der Gefäße und Gefäßbündel, fand demnach erst in der antedevonischen Zeit statt, also im Beginn der zweiten und kleineren Hälfte der organischen Erdgeschichte.
Die Hauptklasse der Farne oder Filicinen wird allgemein in vier verschiedene Klassen eingetheilt, nämlich 1. die Schaftfarne oder Calamophyten, 2. die Laubfarne oder Geopteriden, 3. die Wasserfarne oder Hydropteriden, und 4. die Schuppenfarne oder Lepidophyten. Die bei weitem wichtigste und formenreichste von diesen vier Klassen, welchen den Hauptbestandtheil der paläolithischen Wälder bildete, waren die Laubfarne, und demnächst die Schuppenfarne. Dagegen traten die Schaftfarne schon damals mehr zurück und von den Wasserfarnen wissen wir nicht einmal mit Bestimmtheit, ob sie damals schon lebten. Es muß uns schwer fallen, uns eine Vorstellung von dem ganz eigenthümlichen Charakter jener düsteren paläolithischen Farnwälder zu bilden, in dene der ganze bunte Blumenreichthum unserer gegenwärtigen Flora noch völlig fehlte, und welche noch von keinem Vogel belebt wurden. Von Blumenpflanzen existirten damals nur die beiden niedersten Klassen, die nacksamigen Nadelhölzer und Palmfarne, deren einfache und unscheinbare Blüthen kaum den Namen der Blumen verdienen. Wahrscheinlich sind alle vier Farnklassen als vier getrennte Aeste des Stammbaums zu betrachten, die aus einem gemeinsamen Hauptaste in der Antedevonzeit ihren Ursprung nahmen. Jedoch sind einerseits die niederen Schaftfarne näher mit den Laubfarnen, andrerseits die höheren Schuppenfarne näher mit den Wasserfarnen verwandt, so daß man auch zwei gabelspaltige Aeste oder einen doppelt gabelspaltigen Hauptast als die Stammbasis der ganzen Farnhauptklasse ansehen kann.
Auf der niedersten Organisationsstufe bleibt unter den Farnen die erste Klasse stehen, die Schaftfarne (Calamariae oder Calamophyta). Sie umfaßt drei verschiedene Ordnungen, von denen nur eine noch gegenwärtig lebt, nämlich die Schafthalme (Equisetaceae). Die beiden anderen Ordnungen, die Riesenhalme (Calamiteae)und die Sternblatthalme (Asterophylliteae) sind längst ausgestorben.
Alle Schaftfarne zeichnen sich durch einen hohlen und gegliederten Schaft, Stengel oder Stamm aus, an welchem Aeste und Blätter, wenn sie vorhanden sind, quirlförmig um die Stengelglieder herumstehen. Die hohlen Stengelglieder sind durch Querscheidewände von einander getrennt. Bei den Schafthalmen und Calamiten ist die Oberfläche von längsverlaufenden parallelen Rippen durchzogen, wie bei einer cannulirten Säule, und die Oberhaut enthält so viel Kieselerde, daß sie zum Scheuern und Poliren verwendet werden kann. Bei den Sternblatthalmen oder Asterophyten waren die sternförmig in Quirle gestellten Blätter stärker entwickelt als bei den beiden anderen Ordnungen. In der Gegenwart leben von den Schaftfarnen nur noch die unansehnlichen Schafthalme oder Equisetum-Arten unserer Sümpfe und Moore, welche während der ganzen Primär- und Secundärzeit durch mächtige Bäume aus der Gattung Equisetites vertreten waren.
Zur selben Zeit lebte auch die nächstverwandte Ordnung der Riesenhalme (Calamites), deren starke Stämme gegen 50 Fuß Höhe erreichten. Die Ordnung der Sternblatthalme (Asterophyllites) dagegen enthielt kleinere, zierliche Pflanzen von sehr eigenthümlicher Form, und blieb ausschließlich auf die Primärzeit beschränkt. Die Hauptmasse der Farngruppe bildete zu allen Zeiten die Klasse der eigentlichen Farne im engeren Sinne, der Laubfarne oder Wedelfarne (Filices), auch Landfarne oder Geopteriden genannt, im Gegensatz zu den Wasserfarnen oder Hydropteriden. In der gegenwärtigen Flora unserer gemäßigten Zonen spielt diese Klasse nur eine untergeordnete Rolle, da sie hier meistens nur durch die niedrigen stammlosen Farnkräuter vertreten ist. In der heißen Zone dagegen, namentlich in den feuchten, dampfenden Wäldern der Tropengegenden erhebt sie sich noch heutigentags zur Bildung der hochstämmigen, palmenähnlichen Farnbäume.
Diese schönen Baumfarne der Gegenwart, welche zu den Hauptzierden unserer Gewächshäuser gehören, können uns aber nur eine schwache Vorstellung von den stattlichen und prachtvollen Laubfarnen der Primärzeit geben, deren mächtige Stämme damals dichtgedrängt ganze Wälder zusammensetzten. Man findet diese Stämme namentlich in den Steinkohlenflötzen der Carbonzeit massenhaft ueber einander gehäuft, und dazwischen vortrefflich erhaltene Abdrücke von den zierlichen Wedeln oder Blättern, welche in schirmartig ausgebreitetem Busche den Gipfel des Stammes krönten. Die einfache oder mehrfache Zusammensetzung und Fiederung dieser Wedel, der zierliche Verlauf der verästelten Nerven oder Gefäßbündel in ihrem zarten Laube ist an den Abdrücken der paläolithischen Farnwedel noch so deutlich zu erkennen, wie an den Farnwedeln der Jetztzeit. Bei Vielen sind selbst die Fruchthäufchen, welche auf der Unterfläche der Wedel vertheilt sind, ganz deutlich erhalten. Nach der Steinkohlenzeit nahm das Uebergewicht der Laubfarne bereits ab und schon gegen Ende der Secundärzeit spielten sie eine fast so untergeordnete Rolle wie in der Gegenwart.
Am wenigsten bekannt von allen Farnen ist uns die Geschichte der dritten Klasse, der Wurzelfarne oder Wasserfarne (Rhizocarpeae oder Hydropterides). In ihrem Bau schließen sich diese, im süßen Wasser lebenden Farne einerseits an die Laubfarne, andrerseits an die Schuppenfarne an, sind jedoch den letzteren und dadurch auch den Blumenpflanzen näher verwandt, als die letzteren. Es gehören hierher die wenig bekannten Mosfarne (Salvinia), Kleefarne (Marsilea) und Pillenfarne (Pilularia) in den süßen Gewässern unserer Heimat, ferner die größere schwimmende Azolla der Tropenteiche. Die meisten Wasserfarne sind von zarter Beschaffenheit und deshalb wenig zur Versteinerung geeignet. Daher mag es wohl rühren, daß ihre fossilen Reste so selten sind, und daß die ältesten derselben, die wir kennen, im Jura gefunden wurden. Wahrscheinlich ist aber die Klasse viel älter und hat sich bereits während der paläolithischen Zeit aus anderen Farnen durch Anpassung an das Wasserleben entwickelt.
Die vierte und letzte Farnklasse bilden die Schuppenfarne (Lepidophyta oder Selagines). Sie entwickeln sich höher als alle übrigen Farne und bilden bereits den Uebergang zu den Blumenpflanzen, die sich aus ihnen zunächst hervorgebildet haben. Nächst den Wedelfarnen waren sie am meisten an der Zusammensetzung der paläolithischen Farnwälder betheiligt. Auch diese Klasse enthält, gleichwie die Klasse der Schaftfarne, drei nahe verwandte, aber doch mehrfach verschiedene Ordnungen, von denen nur noch eine am Leben, die beiden anderen aber bereits gegen Ende der Steinkohlenzeit ausgestorben sind. Die heute noch lebenden Schuppenfarne gehören zur Ordnung der Bärlappe (Lycopodiaceae). Es sind meistens kleine und zierliche, mosähnliche Pflänzchen, deren zarter, in vielen Windungen schlangenartig auf dem Boden kriechender und vielverästelter Stengel dicht von schuppenähnlichen und sich deckenden Blättchen eingehüllt ist.
Die zierlichen Lycopodium-Ranken unserer Wälder, welche die Gebirgsreisenden um ihre Hüte winden, werden Ihnen Allen bekannt sein, ebenso die noch zartere Selaginella, welche als sogenanntes "Rankenmos" den Boden unserer Gewächshäuser als dichter Teppich ziert. Die größten Bärlappe der Gegenwart leben auf den Sundainseln und erheben sich dort zu Stämmen von einem halben Fuß Dicke und 25 Fuß Höhe. Aber in der Primärzeit und Secundärzeit waren noch größere Bäume dieser Art weit verbreitet, von denen die ältesten wahrscheinlich zu den Stammeltern der Nadelhölzer gehören (Lycopodites). Die mächtigste Entwickelung erreichte jedoch die Klasse der Schuppenfarne während der Primärzeit nicht in den Bärlappbäumen, sondern in den beiden Ordnungen der Schuppenbäume (Lepidodendreae) und der Siegelbäume (Sigilarieae). Diese beiden Ordnungen treten schon in der Devonzeit mit einzelnen Arten auf, erreichen jedoch ihre massenhafte und erstaunliche Ausbildung erst in der Steinkohlenzeit, und sterben bereits gegen Ende derselben oder in der darauf folgenden Antepermzeit wieder aus. Die Schuppenbäume oder Lepidodendren waren wahrscheinlcih den Bärlappen noch näher verwandt, als die Siegelbäume. Sie erhoben sich zu prachtvollen, unverästelten und gerade aufsteigenden Stämmen, die sich im Gipfel nach Art eines Kronleuchters gabelspaltig in zahlreiche Aeste theilten. Diese trugen eine mächtige Krone von Schuppenblättern und waren gleich dem Stamm in zierlichen Spirallinien von den Narben oder Ansatzstellen der abgefallenen Blätter bedeckt.
Man kennt Schuppenbäume von 40-60 Fuß Lange und 12-15 Fuß Durchmesser am Wurzelende. Einzelne Stämme sollen selbst mehr als hundert Fuß lang sein. Noch viel massenhafter finden sich in der Steinhohle die nicht minder hohen, aber schlankeren Stämme der merkwürdigen Siegelbäume oder Sigillarien angehäuft, die an manchen Orten hauptsächlich die Steinkohlenflötze zusammensetzen. Ihre Wurzelstöcke hat man früher als eine ganz besondere Pflanzenform (Stigmaria beschrieben. Die Siegelbäume sind in vieler Beziehung den Schuppenbäumen sehr ähnlich, weichen jedoch durch ihren anatomischen Bau schon mehrfach von diesen und von den Farnen überhaupt ab, und scheinen einen Uebergang zu den Gymnospermen, insbesondere zu den Palmfarnen oder Cycadeen zu bilden.
Indem wir nun die dichten Farnwälder der Primärzeit verlassen, welche vorzugsweise aus den Laubfarnen, aus den Schuppenbäumen und Siegelbäumen zusammengesetzt sind, treten wir in die nicht minder charakteristischen Nadelwälder der Secundärzeit hinüber. Damit treten wir aber zugleich aus dem Bereiche der blumenlosen Pflanzen oder Kryptogamen in die zweite Hauptabtheilung des Pflanzenreichs, in das Unterreich der Blumenpflanzen oder Phanerogamen hinein. Diese formenreiche Abtheilung, welche die Hauptmasse der jetzt lebenden Pflanzenwelt, und namentlich die große Mehrzahl der landbewohnenden Pflanzen enthält, ist jedenfalls viel jüngeren Alters als die Abtheilung der Kryptogamen. Denn sie kann erst im Laufe des paläolitischen Zeitalters aus dieser letzteren sich entwickelt haben. Mit voller Gewißheit können wir behaupten, daß während des ganzen archolithischen Zeitalters, also während der ersten und längeren Hälfte der organischen Erdgeschichte, noch gar keine Blumenpflanzen existirten, und daß sie sich erst während der Primärzeit aus farnartigen Kryptogamen entwickelt haben. Die anatomische und embryologische Verwandtschaft der Phanerogamen mit diesen letzteren ist so innig, daß wir daraus mit Sicherheit auch auf ihren genealogischen Zusammenhang, ihre wirkliche Blutsverwandtschaft schließen können. Die Blumenpflanzen können unmittelbar weder aus Thalluspflanzen noch aus Mosen, sondern nur aus Farnen oder Filicinen entstanden sein. Höchst wahrscheinlich sind die Schuppenfarne oder Lepidophyten, und zwar Bärlapppflanzen oder Lycopodiaceen, welcher der heutigen Selaginella sehr nahe verwandt waren, die unmittelbaren Vorfahren der Phanerogamen.
Schon seit langer Zeit hat man auf Grund des inneren anatomischen Baues und der embryologischen Entwickelung das Unterreich der Phanerogamen in zwei große Hauptklassen eingetheilt, in die Nacktsamigen oder Gymnospermen und in die Decksamigen oder Angiospermen. Diese letzteren sind in jeder Beziehung vollkommener und höher organisiert als die ersteren, und haben sich erst später, im Laufe der Secundärzeit, aus diesen entwickelt. Die Gymnospermen bilden sowohl anatomisch als embryologisch die vermittelnde Uebergangsgruppe von den Farmen zu den Angiospermen.
Die niedere, unvollkommenere und ältere von den beiden Hauptklassen der Blumenpflanzen, die der Nacktsamigen (Gymnospermae) erreichte ihre mannichfaltige Ausbildung und ihre weiteste Verbreitung während der mesolithischen oder Secundärzeit. Sie ist für dieses Zeitalter nicht minder charakteristisch, wie die Farngruppe für das vorhergehende primäre, und wie die Angiospermengruppen für das nachfolgende tertiäre Zeitalter. Wir konnten daher die Secundärzeit auch als den Zeitraum der Gymnospermen, oder nach ihren bedeutendsten Vertretern als das Zeitalter der Nadelhölzer bezeichnen. Von den beiden Klassen, in welche die Gymnospermen zerfallen, den Nadelhölzern und Palmfarnen, ist die erstere am stärksten in der Triaszeit, die letztere in der Jurazeit entwickelt. Jedoch fällt die Entstehung der ganzen Hauptklasse schon in eine frühere Zeit. Wir finden versteinerte Reste von beiden Klassen derselben bereits in der Steinkohle vor, und müssen daraus schließen, daß der Uebergang von Schuppenfarnen in Gymnospermen bereits während der Steinkohlenzeit, oder vielleicht schon vorher, in der antecarbonischen oder in der devonischen Zeit erfolgt ist. Immerhin spielen die Nacktsamigen während der ganzen folgenden Primärzeit nur eine sehr untergeordnete Rolle und gewinnen die Herrschaft über die Farne erst im Beginn der Secundärzeit.
Von den beiden Klassen der Gymnospermen steht diejenige der Palmfarne oder Zamien (Cycadeae) auf der niedersten Stufe und schließt sich, wie schon der Name sagt, unmittelbar an die Farne an, so daß sie selbst von manchen Botanikern wirklich mit dieser Gruppe im Systeme vereinigt werden. In der äußeren Gestalt gleichen sie sowohl den Palmen aus den Farnbäumen oder baumartigen Laubfarnen und tragen eine aus Fiederblättern zusammengesetzte Krone, welche entweder auf einem dicken niedrigen Strunke oder auf einem schlanken, einfachen, säulenförmigen Stamme sitzt. In der Gegenwart ist diese einst formenreiche Klasse nur noch durch wenige, in der heißen Zone lebende Formen dürftig vertreten, durch die niedrigen Zapfenfarme (Zamia), die dickstämmigen Brodfarne (Encephalartos), und die schlankstämmigen Rollfarne (Cycas). Man findet sie häufig in unseren Treibhäusern, wo sie gewöhnlich mit Palmen verwechselt werden. Eine viel größere Formenmannichfaltigkeit als die lebenden, bieten uns die ausgestorbenen und versteinerten Zapfenfarne, welche namentlich in der Mitte der Secundärzeit, während der Juraperiode in größter Masse auftraten und damals vorzugsweise den Charakter der Wälder bestimmten. Gymnospermen, welche diesen Cycadeen nächstverwandt und vielleicht nicht von ihnen zu trennen waren, erzeugten während der älteren oder mittleren Secundärzeit die Hauptklasse der Angiospermen.
In größerer Formenmannichfaltigkeit als die Klasse der Palmfarne hat sich bis auf unsere Zeit der andere Zweig der Gymnospermengruppe erhalten, die Klasse der Nadelhölzer oder Zapfenbäume (Coniferae). Noch gegenwärtig spielen die dazu gehörigen Cypressen, Wachholder und Lebensbäume (Thuja), die Taxus und Ginkobäume (Salisburya), die Araucarien und Cedern, vor allen aber formenreiche Gattung Pinus mit ihren zahlreichen und bedeutenden Arten, den verschiedenen Kiefern, Pinien, Tannen, Fichten, Lärchen u. s. w. in den verschiedensten Gegenden der Erde eine sehr bedeutende Rolle, und setzen ausgedehnte Waldgebiete fast allein zusammen. Doch erscheint diese Entwickelung der Nadelhölzer schwach im Vergleiche zu der ganz überwiegenden Herrschaft, welche sich diese Klasse während der älteren Secundärzeit, in der Triasperiode, über die übrigen Pflanzen erworben hatte. Damals bildeten mächtige Zapfenbäume in verhältnißmäßig wenigen Gattungen und Arten, aber in ungeheuren Massen von Individuen beisammen stehend, den Hauptbestandtheil der mesolithischen Wälder. Sie rechtferigen die Benennung der Secundärzeit als des "Zeitalters der Nadelwälder", obwohl die Coniferen schon in der Jurazeit von den Cycadeen überflügelt wurden.
Aus den Nadelwäldern der mesolithischen oder Secundärzeit treten wir in die Laubwälder der cenolithischen oder Tertiärzeit hinüber und gelangen dadurch zur Betrachtung der sechsten und letzten Hauptklasse des Pflanzenreichs, der Decksamigen (Angiospermae). Wie schon vorher bemerkt, hat sich diese zweite Hauptklasse der Blumenpflanzen erst viel später als die Nacktsamigen, und zwar aus einem Zweige dieser letzteren entwickelt. Die ersten sicheren und unzweifelhaften Versteinerungen von Decksamigen finden wir in den Schichten des Kreidesystems, und zwar kommen hier neben einander Reste von den beiden Klassen vor, in welche man die Hauptklasse der Angiospermen allgemein eintheilt, nämlich Einkeimblättrige oder Monocotylen und Zweikeimblättrige oder Dicotylen. Die letzteren sind jedenfalls nicht älter als die Kreidezeit oder höchstens die Antecretazeit. Dagegen sind die ersteren möglicherweise auch schon früher vorhanden gewesen. Wir kennen nämlich eine Anzahl von zweifelhaften und nicht sicher bestimmbaren fossilen Pflanzenresten aus der Jurazeit und aus der Triaszeit, welche von manchen Botanikern bereits für Monocotylen, von anderen dagegen für Gymnospermen gehalten werden. Selbst in den Steinkohlenschichten glaubte man Monocotylenreste gefunden zu haben, die sich aber neuerdings als Ueberbleibsel entweder von Nacktsamigen oder von Farnen herausgestellt haben. Demnach scheint es jetzt sicher zu sein, daß die Klasse der Decksamigen erst während der Secundärzeit, und zwar aus den Cycadeen oder diesen nächstverwandten Nacktsamigen entstanden ist. Was die beiden Klassen der Decksamigen betrifft, Monocotylen und Dicotylen, so haben sich entweder beide Zweige aus einem gemeinsamen Stammaste, oder die Dicotylen erst später aus den Monocotylen entwickelt. Jedenfalls stehen in anatomischer Beziehung die letzteren auf einer tieferen und unvollkommeneren Stufe als die ersteren.
Die Klasse der Einkeimblättrigen oder Einsamenlappigen (Monocotylae oder Monocotyledones, auch Endogenae genannt) umfaßt diejenigen Blumenpflanzen, deren Samen nur ein einziges Keimblatt oder einen sogenannten Samenlappen (Cotyledon) besitzt. Jeder Blattkreis ihrer Blume enthält in der großen Mehrzahl der Fälle drei Blätter, und es ist sehr wahrscheinlich, daß die gemeinsame Mutterpflanze aller Monocotylen eine regelmäßige und dreizählige Blüthe besaß. Die Blätter sind meistens einfach, von einfachen, graden Gefäßbündeln oder sogenannten "Nerven" durchzogen. Zu dieser Klasse gehören die umfangreichen Familien der Binsen und Gräser, Lilien, Schwertlilien, Orchideen und Dioscoreen, ferner eine Anzahl einheimscher Wasserpflanzen, die Wasserlinsen, Rohrkolben, Seegräser u. s. w. und endlich die prachtvollen, höchst entwickelten Familien der Aroideen und Pandaneen, der Bananen und Palmen. Im Ganzen ist die Monocotylenklasse trotz aller Formenmannichfaltigkeit, die sie in der Tertiärzeit und in der Gegenwart entwickelt hat, viel einförmiger organisirt, als die Dicotylenklasse, und auch ihre geschichtliche Entwickelung bietet ein viel geringeres Interesse. Da ihre versteinerten Reste meistens schwer zu erkennen sind, so bleibt die Frage vorläufig offen, in welchem der drei großen secundären Zeiträume, Trias-, Jura- oder Kreidezeit, die Monocotylen aus den Cycadeen entstanden sind. Jedenfalls existiren sie in der Kreidezeit schon eben so sicher wie die Dicotylen.
Viel größeres historisches und anatomisches Interesse bietet in der Entwickelung ihrer untergeordneten Gruppen die zweite Klasse der Decksamigen, die Zweikeimblättrigen oder Zweisamenlappigen (Dicotylae oder Dicotyledones, auch Exogenae benannt). Die Blumenpflanzen dieser Klasse besitzen, wie ihr Name sagt, gewöhnlich zwei Samenlappen oder Keimblätter (Cotyledonen). Die Grundzahl in der Zusammensetzung ihrer Blüthe ist gewöhnlich nicht drei, wie bei den meisten Monocotylen, sondern vier oder fünf, oder ein Vielfaches davon. Ferner sind ihre Blätter gewöhnlich höher differenzirt und mehr zusammengesetzt, als die der Monocotylen, und von gekrümmten, verästelten Gefäßbündeln oder "Adern" durchzogen. Zu dieser Klasse gehören die meisten Laubbäume, und da dieselbe in der Tertiärzeit schon ebenso wie in der Gegenwart das Uebergewicht über die Gymnospermen und Farne gewann, so konnten wir das cenolithische Zeitalter auch als das der Laubwälder bezeichnen.
Obwohl die Mehrzahl der Dicotylen zu den höchsten und vollkommensten Pflanzen gehört, so schließt sich doch die niederste Abtheilung derselben unmittelbar an die Monocotylen an und stimmt mit diesen namentlich darin überein, daß in ihrer Blüthe Kelch und Blumenkrone noch nicht gesondert sind. Man nennt sie daher Kelchblüthige (Monochlamydeae oder Apetalae). Diese Unterklasse hat sich zunächst entweder aus den Monocotylen oder in Zusammenhang mit diesen aus den Gymnospermen entwickelt. Es gehören dahin die meisten kätzchentragenden Laubbäume, die Birken und Erlen, Weiden und Pappeln, Buchen und Eichen, ferner die nesselartigen Pflanzen, Nesseln, Hanf und Hopfen, Feigen, Maulbeeren und Rüstern, endlich die Wolfsmilchartigen, Amaranthartigen, Lorberartigen u. s. w. Neben den Kelchblüthigen lebte aber in der Kreidezeit auch schon die zweite und vollkommenere Unterklasse der Dicotylen, die Gruppe der Kronenblüthigen (Dichlamydeae oder Corolliflorae). Diese entstanden aus den Kelchblüthigen dadurch, daß sich die einfache Blüthenhülle der letzteren in Kelch und Krone differenzirte. Die Unterklasse der Kronenblüthigen zerfällt wiederum in zwei große Hauptabtheilungen oder Legionen, deren jede eine große Menge von verschiedenen Ordnungen, Familien, Gattungen und Arten enthält. Die erste Legion führt den Namen der Sternblüthigen oder Diapetalen, die zweite den Namen der Glockenblüthigen oder Gamopetalen.
Die tiefer stehende und unvollkommenere von den beiden Legionen der Kronenblüthigen sind die Sternblüthigen (Diapetalae, auch Polypetalae oder Dialypetalae genannt). Hierher gehören die umfangreichen Familien der Doldenblüthigen oder Umbelliferen, der Kreuzblüthigen oder Cruciferen, ferner die Ranunculaceen und Crassulaceen, Wasserrosen und Cistrosen, Malven und Geranien, und neben vielen anderen namentlich noch die großen Antheilungen der Rosenblüthigen, (welche außer den Rosen die meisten unserer Obstbäume umfassen) und der Schmetterlingsblüthigen, (welche unter anderen die Wicken, Bohnen, Klee, Ginster, Akacien und Mimosen enthalten). Bei allen diesen Diapetalen bleiben die Blumenblätter getrennt und verwachsen nicht mit einander, wie es bei den Gamopetalen der Fall ist. Die letzteren haben sich erst in der Tertiärzeit aus den Diapetalen entwickelt, während diese schon in der Kreidezeit neben den Kelchblüthigen auftraten.
Die höchste und vollkommenste Gruppe des Pflanzenreichs bildet die zweite Abtheilung der Kronenblüthigen, die Legion der Glockenblüthigen (Gamopetalae, auch Monopetalae oder Sympetalae genannt). Hier verwachsen die Blumenblätter, welche bei den übrigen Blumenpflanzen meistens ganz getrennt bleiben, regelmäßig zu einer mehr oder weniger glocken- trichter- oder röhrenförmigen Krone. Es gehören hierher unter anderen die Glockenblumen und Winden, Primeln und Haidekräuter, Gentiane und Gaisblatt, ferner die Familie der Oelbaumartigen, Oelbaum, Liguster, Flieder und Esche, und endlich neben vielen anderen Familien die umfangreichen Abtheilungen der Lippenblüthigen (Labiaten) und der Zusammengesetztblüthigen (Compositen). In diesen letzteren erreicht die Differenzirung und Vervollkommnung der Phanerogamenblüthe ihren höchsten Grad, und wir müssen sie daher als die Vollkommensten von allen an die Spitze des Pflanzenreichs stellen. Dem entsprechend tritt die Legion der Glockenblüthigen oder Gamopetalen am spätesten von allen Hauptgruppen des Pflanzenreichs in der organischen Erdgeschichte auf, nämlich erst in der cenolithischen oder Tertiärzeit. Selbst in der älteren Tertiärzeit ist sie noch sehr selten, nimmt erst in der mittleren langsam zu und erreicht erst in der neueren Tertiärzeit und in der Quartärzeit ihre volle Ausbildung.
Wenn Sie nun, in der Gegenwart angelangt, nochmals die ganze geschichtliche Entwickelung des Pflanzenreichs überblicken, so werden Sie nicht umhin können, darin lediglich eine großartige Bestätigung der Descendenztheorie zu erblicken. Die beiden großen Grundgesetze der organischen Entwickelung, die wir als die nothwendigen Folgen der natürlichen Züchtung im Kampf um's Dasein nachgewiesen haben, die Gesetze der Differenzirung und der Vervollkommnung, machen sich in der Entwickelung der größeren und kleineren Gruppen des natürlichen Pflanzensystems überall geltend. In jeder größeren und kleineren Periode der organischen Erdgeschichte nimmt das Pflanzenreich sowohl an Mannichfaltigkeit, als an Vollkommenheit zu, wie Ihnen schon ein Blick auf Taf. II deutlich zeigt.
Während der ganzen langen Primordialzeit existirte nur die niederste und unvollkommenste Hauptklasse der Tange. Zu diesen gesellten sich in der Primärzeit die höheren und vollkommeneren Kryptogamen, insbesondere die Hauptklasse der Farne. Schon während der Steinkohlenzeit beginnen sich aus diesen die Phanerogamen zu entwickeln, anfänglich jedoch nur durch die niedere Hauptklasse der Nacktsamigen oder Gymnospermen repräsentirt. Erst während der Secundärzeit geht aus diesen die höhere Hauptklasse der Decksamigen oder Angiospermen hervor. Auch von diesen sind anfänglich nur die niederen, kronenlosen Gruppen, die Monocotylen, dann die Apetalen vorhanden. Erst während der Kreidezeit entwickelten sich aus letzteren die höheren Kronenblüthigen. Aber auch diese höchste Abtheilung ist in der Kreidezeit nur durch die tiefer stehenden Sternblüthigen oder Diapetalen vertreten, und ganz zuletzt erst, in der Tertiärzeit, gehen aus diesen die höher stehenden Glockenblüthigen oder Gamopetalen hervor, die vollkommendsten von allen Blumenpflanzen. So erhob sich in jedem jüngeren Abschnitt der organischen Erdgeschichte das Pflanzenreich stufenweise zu einem höheren Grade der Vollkommenheit und der Mannichfaltigkeit.
Ich habe Ihnen in dieser systematischen Uebersicht über die historische Entwickelung des Pflanzenreichs dasselbe als eine einzige Gruppe von blutsverwandten Organismen dargestellt, wie es auch der Stammbaum auf Taf. II ausdrückt. Mir scheint diese einstämmige oder monophyletische Anschauung vom Ursprung des Pflanzenreichs die naturgemäßere zu sein. Damit will ich jedoch nicht sagen, daß dieselbe nothwendig die allein richtige ist. Es läßt sich auch denken, daß das Pflanzenreich aus mehreren selbstständigen Stämmen oder Phylen zusammengesetzt ist, deren jeder aus einer einzigen archigonen (d. d. durch Urzeugung entstandenen) Monerenart hervorgegangen ist. Eine Vorstellung von dieser vielstämmigen oder polyphyletischen Descendenzhypothese mag Ihnen nachstehende Tabelle geben. Kaum zweifelhaft ist es, daß auch in diesem Falle die ganze Masse der Stockpflanzen oder Cormophyten (sowohl Phanerogamen als Prothallophyten) als Blutsverwandte eines einzigen Stammes aufzufassen sind.
Denn die genealogische Stufenleiter von den Mosen zu den Farnen, von diesen zu den Nacktsamigen, und von letzteren zu den Decksamigen, ebenso innerhalb der letzten Gruppe die Stufenleiter von den Kelchblüthigen (Monocolylen und Apetalen) zu den Kronenblüthigen (Diapetalen und Gamopetalen) wird zu klar durch das übereinstimmende Zeugniß der vergleichenden Anatomie, Ontogenie und Paläntologie bewiesen, als daß man an einer Blutsverwandtschaft aller dieser Cormophyten zweifeln könnte. Dagegen ist es wohl möglich, daß die verschiedenen Gruppen der Thallophyten von mehreren (und vielleicht von zahlreichen) verschiedenen Moneren, die durch wiederholte Urzeugungsakte entstanden, abstammen. Als ein ganz selbstständiges Phylum ließe sich z. B. auffassen die Klasse der Fucioideen, als ein zweites die Klasse der Florideen, als ein drittes die Klasser der Flechten. Die drei Klassen der Pilze, Grüntange und Urtange sind vielleicht aus zahlreichen, ganz unabhängigen Phylen zusammengesetzt, und dann würde ein einzelnes Phylum der Grüntange den ganzen Stamm der Cormophyten erzeugt haben. Es ist möglich, daß zukünftige Untersuchungen uns über diese sehr dunkle und schwierige Frage noch etwas aufklären werden. Uebrigens ist dieselbe nur von sehr untergeordnetem Intersse, da unsere monophyletische Anschauung von dem einheitlichen Ursprunge der bei weitem größten und wichtigsten Pflanzengruppe, der Cormophyten, dadurch gar nicht berührt wird. (Vergl. Gen. Morph. II, Taf. II, S. XXXI und 406). { Abbildung Seite 382 }
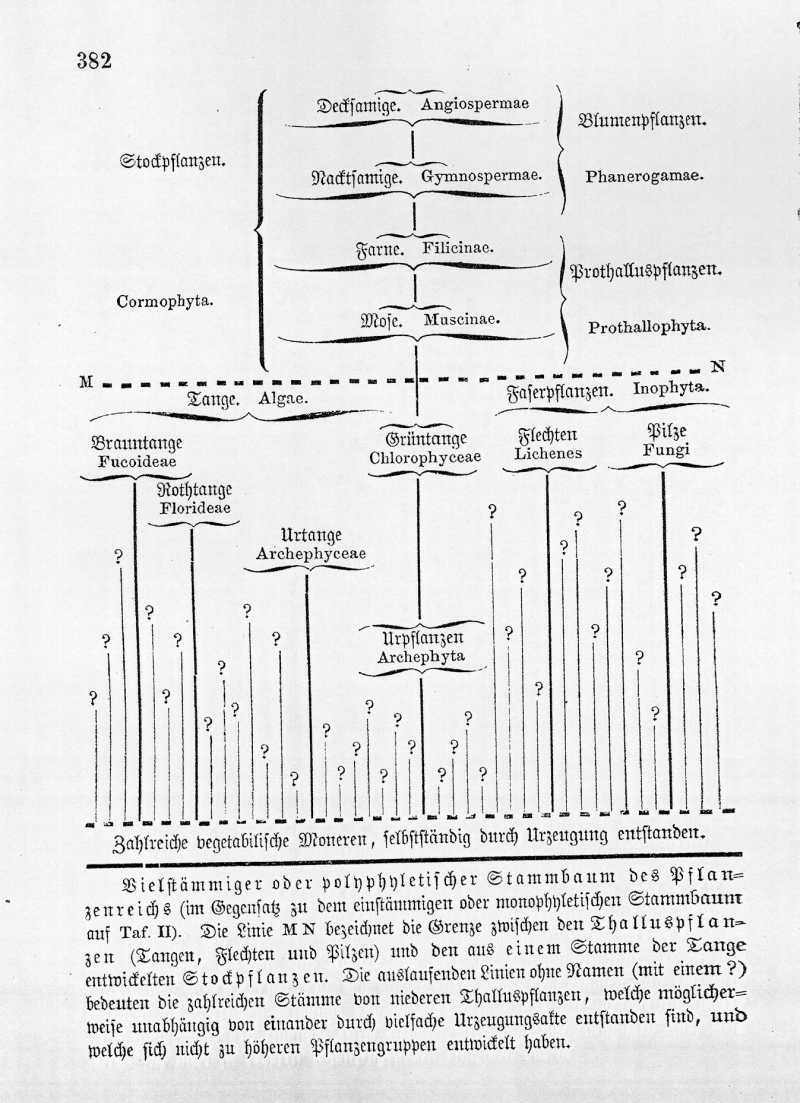
Vielstämmiger oder polyphyletischer Stammbaum des Pflanzenreichs (im Gegensatz zu dem einstämmigen oder monophyletischen Stammbaum auf Taf. II). Die Linie M N bezeichnet die Grenze zwischen den Thalluspflanzen (Tangen, Flechten und Pilzen) und den aus einem Stamme der Tange entwickelten Stockpflanzen. Die auslaufenden Linien ohne Namen (mit einem ?) bedeuten die zahlreichen Stämme von niederen Thalluspflanzen, welche möglicherweise unabhängig von einander durch vielfache Zeugungsakte entstanden sind, und welche sich nicht zu höheren Pflanzengruppen entwickelt haben.
Stammbaum und Geschichte des Protistenreiches

 Stammbaum und Geschichte des Thierreichs
Stammbaum und Geschichte des Thierreichs
Renaissance Forum - Rätselverzeichnis - Wie alles begann ... - Zufallsrätsel - erstes Rätsel
Galerie - A - B - C - D - E - F- G - H - I - J - K - L - M - N - O - P - Q - R - S - T - U - V - W - X - Y - Z |
Jacob Burckhardt - Die Kultur der Renaissance in Italien. Ein Versuch
Leonardo da Vinci Wissenschaftler - Erfinder - Künstler
Venedig - Eine Liebeserklärung an eine Stadt
William Shakespeare animiert (wahrscheinlich oder zufällig...2/3 zu 1/3) William Turner
Philosophie für Schnelldenker - Besinnliche Philosophie
Startseite - © Copyright 2004- - Susanne Albers - Kiehlufer 125 - D 12059 Berlin